On the Bioprotective Effectiveness of Trehalose: Structural Properties of the Disaccharide Aqueous Mixtures
The main purpose of the present work is to review some structural properties of trehalose-water mixtures to highlight the role played by the sugar in...
The main purpose of the present work is to review some structural properties of trehalose-water mixtures to highlight the role played by the sugar in the modification of the original hydrogen bond network of pure water in order to get some insight into the molecular mechanisms of bioprotection that characterize trehalose.
The complex world of living beings is characterized by mechanisms and laws governing the life, the equilibrium and the interactions among different organisms. In this frame fundamental is the role played by some molecules, such as trehalose, which show a bioprotective action on living organisms.
Trehalose is a non-reducing disaccharide of glucose widely distributed in both prokaryotic and eukaryotic organisms, that, very surprisingly, show the ability of adaptation to extreme environmental stress conditions.
Living beings capable of surviving extreme conditions enter into a state of suspended animation, called cryptobiosis. The word cryptobiosis comes from the Greek kryptos, or hidden, and bios, or living, so it means "hidden life."
According to Keilin (1), cryptobiosis is defined as "the state of an organism when it shows no visible signs of life and when its metabolic activity becomes hardly measurable, or comes to a standstill".
Cryptobiosis occurs in a wide variety of organisms, such as bacteria, fungi, seeds of higher plants, and animals (including tardigrades and eggs of some crustacean (1-2)).
One of the most impressive examples of the biological role of trehalose is represented by tardigrades (water bears), that, thanks to the disaccharide synthetized in cryptobiotic state, are able to survive in extreme conditions, such as temperatures near absolute zero and above the boiling point of water (155°C), pressure of 6000atm, a hard vacuum (as in outer space), high doses radiation, X-rays, nitric acid, hydrochloric acid, carbon dioxide and carbon monoxide. They can live more than 100 years, alternating periods of "true" active life and of "suspended" life (3).
There are different theories to explain the biological role of trehalose (and also sucrose and maltose). Green and Angell (4,5) suggest that the higher value of the glass transition temperature of trehalose and its mixtures with water, in comparison with that of the other disaccharides, is the only reason for its superior bioprotectant effectiveness: this would imply that, at a given temperature, the glass transition for trehalose mixtures always occurs at a higher water content. As a matter of fact such a hypothesis alone is not entirely satisfactory because of the many counter-examples. For example dextran, a linear polysaccharide with α(1-6) glycosidic links, presents even a higher Tg value but does not show a comparable bioprotective action.
[inline: 1= Figure 1: INS spectra of disaccharides] Figure 1: INS spectra of trehalose/2H2O mixture with the MNPS correction. In the insert a zoom of the bending region for disaccharides mixtures shows the more marked "cryptocrystalline" character of trehalose
Crowe and co-workers (6,7) formulated the hypothesis that a direct interaction between the sugars and the object of protection occurs.
Figure1_INS_spectra_of_disaccharides.jpg - [640 x 480 pixel]
More specifically their "water replacement hypothesis" justifies the trehalose protective function with the existence of direct hydrogen bonding of trehalose with the polar head groups of the lipids as water does.
This hypothesis was supported by some simulation studies (8), which argue that the structure of trehalose is perfectly adaptable to the tetrahedral coordination of pure water, whose structural and dynamical properties are not significantly affected by trehalose.
Cordone (9) and Cottone (10) by complementary techniques showed that the mean square displacements of protein atoms are reduced by the presence of trehalose and that sugar binds to the protein through single hydrogen bonds giving rise to water molecules localized at the protein interface; in other words sugar matrices lock the surface of the protein hindering large amplitude solvent coupled protein motions.
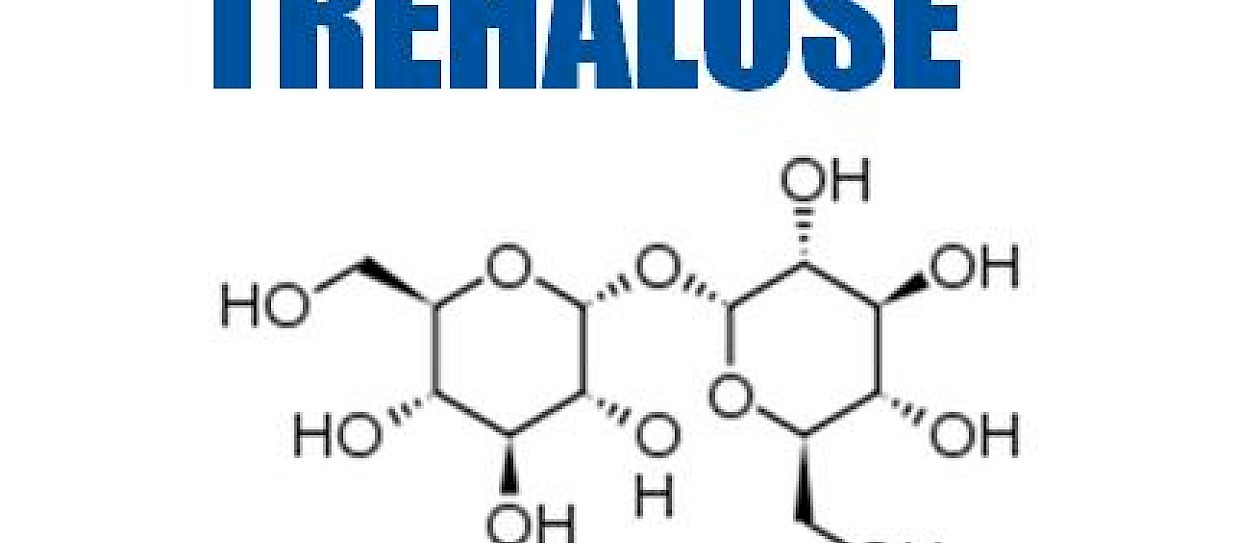
From a general point of view, trehalose, maltose and sucrose have the same chemical formula (C12H22O11) (Molecular Weight Mw=342.3), but different structures.
More precisely, α,α-trehalose is a disaccharide of glucose (α-D-glucopyranosil α-D-glucopyranoside) constituted by two pyranose (six-membered) rings in the same α configuration, linked by a glycosidic bond between the chiral carbon atoms C1 of the two rings.
It is the only isomer of trehalose found in nature, probably first isolated, from the ergot of rye, in 1832 by Wiggers. β,β-trehalose (isotrehalose), the other known trehalose isomers, have not yet been discovered in nature.
Maltose (4-O-α-D-glucopyranosil-D-glucose) is also constituted by two pyranose rings of glucose in the α configuration, but the oxygen bridge links the two carbon atoms C1 and C4 of the two rings.
Finally, sucrose (α-D-glucopyranosil β-D-fructofuranoside) is constituted by a glucose ring (pyranose) in the α configuration and a fructose ring (furanose) in the β configuration; the α and β structures of the same monosaccharide differ only in the orientation of the OH groups at some carbon atom in the ring itself (mutarotation equilibria).
Vibrational spectra of disaccharides/H2O mixtures at T=27 K have been collected by the indirect geometry time-of-flight spectrometer TOSCA at the ISIS Pulse Neutron Facility (RAL, UK) in the energy range 0÷4000 cm-1 (0÷500 meV) with the best results below 2000 cm-1 (250 meV), (11,12).
By this spectrometer both the collective excitations region at low frequency and the localized excitations region at the intermediate and higher frequency region can be covered (13).
TOSCA’s resolution (in the whole energy range of interest, the instrument provides very good energy resolution, about ΔE/E≈1.5÷2%, which is comparable with optical techniques such as IR and Raman spectroscopies), combined with the high intensity of the ISIS source, allows to perform the study of the dynamics with high accuracy, reducing the background noise/signal ratio down to negligible for strongly scattering samples.
Inelastic Neutron Scattering provides a suitable probe for studies of vibrational dynamics in condensed materials, furnishing vibrational spectra which are different from Infrared (IR) and Raman spectroscopies due to various factors: i) for neutrons, no selection rule holds, because of the nuclear nature of the atomic scattering potential; ii) even if, as for IR and Raman spectra, in INS the frequencies are determined by the normal modes of vibration, the intensities depend, besides on the neutron scattering cross-section and on the moment transfer, on the amplitudes of vibrations; as a consequence, the band intensities in neutron spectroscopy allow to obtain precious information about the atomic displacements; iii) in INS spectra the low wavenumber vibrations give more intense bands than the high wavenumber ones; iv) finally, due to the neutron sensitiveness to hydrogen vibrations, INS is particularly important in the study of H bonded systems.
From a formal point of view, the theoretical treatment of INS results moves by the well known expression for the dynamical structure factor:
[inline: 3= Expression for the dynamical structure factor] Expression for the dynamical structure factor
Performing the Fourier transform from the time to the frequency domain it results that the time independent terms will include contributions from all frequencies whilst time dependent exponential terms will be specific to particular frequencies.
The usual one-phonon expression for high ω and low T values reduces to:
[inline: 4= one-phonon expression] one-phonon expression
The observed intensity S(Q,ω) of a band at a frequency ω is function of the momentum transferred Q (determined by spectrometer) and the vibrational amplitude of the hydrogen atoms that can be extracted (14-16). The multiphonon neutron scattering contribution (MPNS) has been evaluated by means of an iterative technique based on the assumption that the one-phonon term describes the INS spectrum at the lower energy transfers and on the calculation of the first MPNS term for the whole energy region from this first step. The result is a new one-phonon spectrum that will allow to get with successive iterative calculations MPNS terms until good convergence is obtained (14,15).
INS spectra of ice can be subdivided into two different regions: i) the region of the H2O intermolecular vibrations up to ˜960 cm-1 , in which one distinguishes a "translational" part up to ˜400 cm-1 and a "librational" part, ii) the region of the H2O intramolecular vibrations up to &tile;3600 cm-1, that presents the bending modes at ˜1600 cm-1 and the stretching modes at ˜3360 cm-1.
As far as the INS spectra of disaccharides/H2O mixtures are concerned, Figure 1 shows, within the spectral frame of ice, the spectrum of trehalose/H2O mixture at a concentration value corresponding to 2 H2O molecules for each disaccharide molecule together with the MPNS correction. As it can be seen, this correction does not affect significantly the translational modes region at low frequency, but increases at larger energy transfer.
More specifically, taking into account the different spectral regions, the following features can be observed:
i) 0÷200 cm-1 region
This spectral region concerns with low energy collective modes. No significant difference is observed for the three disaccharides/H2O mixtures: as an example, the peak at ˜54 cm-1 in the ice spectrum, associated with the distortion of the local structure due to the topological disorder, is equivalently depressed by the three disaccharides.
ii) 200÷1060 cm-1 region
As already mentioned, in this region for ice two different contributions to the intermolecular modes can be distinguished: translational and librational modes, respectively. The spectra of the disaccharides/H2O mixtures in the region 200÷400 cm-1 are all dominated by ice. Important differences for the three disaccharides are observed in the 400÷1060 cm-1 region.
It was found that the characteristic value for the librational band in the INS spectrum for different ice forms is the position of its low-energy cut-off (14-16). Assuming the moment of inertia for water molecules in the ices is about the same, the observed shifts of the cut-off position indicate that the transverse forces between the water molecules are of different intensity for the different ice forms.
We observe that in the ice spectrum the position of this cut-off is at ˜ 544 cm-1, whereas for the investigated mixtures we find for its position the value of ˜ 547 cm-1 for trehalose and maltose and of ˜ 544 cm-1 for sucrose. This circumstance clearly indicates that trehalose and maltose influence more than sucrose the interactions among different H2O molecules. On the other hand, the intensity increase at the cut-off for ice is ˜3.61, whereas for trehalose, maltose and sucrose ˜1.84, ˜1.91 and 2.03, respectively.
In trehalose vibrational spectrum obtained by simulation (17) we note that the peak attributable to the C-O stretching mode at ˜1000 cm-1 does not appear, or at least is not detectable, indicating that this kind of modes in the rings is constrained because of the presence of hydrogen bonds with water. In general terms, however, below 960 cm-1 it is not possible to assign a well defined character to all the bands (15). The stretching of the C-O bonds occurs in a range between 970 and 1100 cm-1 , while it is difficult to assign a well-defined character to most of the modes below 950 cm-1: only the oscillations of the OH groups around the minimum of the C-C-O-H torsional potential are well localized and fairly easy to identify. These modes have energy between 420 and 570 cm-1, with the exception of the OH group giving rise to the intramolecular hydrogen bond, whose torsional mode has energy of 700 cm-1 (17).
These findings suggest that all the disaccharides modify significantly the spectral features of ice in this spectral region.
In particular trehalose shows the smaller value of its intensity increase and the larger shift of its position, this circumstance indicating that it affects more than the other disaccharides the intermolecular interaction forces, and the arrangements of the H2O molecules.
On the other hand, sucrose has no effect on the position, but it changes in intensity, showing, however, a small effect.
Since the attention is focused on the disaccharide-H2O interaction, all these results justify the greater effectiveness of trehalose as structure-breaker of the hydrogen bonded network of ice.
iii) 1060÷2000 cm-1 region
The intensity profile of trehalose/H2O mixture in this spectral region which corresponds to the range of the bending vibrational modes of ice appears more "structured" in comparison with the other disaccharides and in particular with sucrose, this latter showing a more smooth trend.
In Figure 1 a comparison among trehalose, maltose and sucrose/H2O mixtures in the spectral bending region is reported. We retain these comparisons are necessary for pointing out the strong interaction of the H2O molecules with disaccharides, in particular with trehalose.
Following the simulation results (17) the stretching of the C-O bonds occurs in the range between 960 and 1104 cm-1, whereas the band of the hybridized H-C-H, C-C-H and C-O-H bending modes, which in trehalose vibrational spectrum occurs in the range 1175÷1433 cm-1, is present between 1160 and 1440 cm-1.
A striking feature characterizing the simulation results consists in the evaluation of the conformational freedom in the trehalose molecule: the results show that a relatively low energy is required for the torsions around the oxygen linking the two rings to occur, whereas the bending of the same angle is much less likely, being energetically unfavourite.
When the interaction with water is introduced through the simulation of the trehalose crystal with two trehalose monohydrate molecules in the unit cell, apart from some expected distortions in the O-H bonds of water molecules, the most apparent difference between the vibrational spectra of the crystal and of the molecules concerns the modes localized on the OH groups. The C-H stretching band is relatively unchanged in frequency, but results broader. At 1630 cm-1 the isolated peak of the H-O-H bending of water appears. Below this frequency, the differences between the vibrational frequencies of the crystal and of the molecule are quantitatively small, and difficult to identify.
The most apparent effect is an increase of the energy for the OH rotations around the minimum of the torsional potential, due to the formation of the intermolecular hydrogen bonds (17).
It is evident that trehalose shows more distinctly, in respect to the other disaccharides, the characteristic peaks typical of the crystalline state above described. In other terms the trehalose/H2O complex shows a more "crystalline" behaviour, namely a locally more ordered structure. Concluding, trehalose is capable to destroy the tetrahedral network of water and it forms with H2O a unique entity, having a "cryptocrystalline" character, i. e. a nanoscopic crystalline nature, where biostructures can be protected. In our opinion this circumstance is relevant because it implies a better cryobiotic effect.
References:
-
Keilin FRS. The problem of anabiosis or latent life: history and current concept. Sci. Progr. 1959 41:577-591.
-
Hinton HE. Cryptobiosis in the larvae of Polypedilum vanderplanki Hint. (Chironomidae). J. Ins. Physiol. 1960 5:286-300.
-
Miller WR. Tardigrades: Bears of the Moss. Kansas School Naturalist. 1997 43.
-
Green JL, Angell CA. Phase Relations and Vitrification in Saccharide-Water Solutions and the Trehalose Anomaly. J. Phys. Chem. B 1989 93:2880-2882.
-
Angell CA. Glassforming Liquids with Microscopic to Macroscopic Two-State Complexity. Prog. Theoret. Phys. 1997 126:1-7.
-
Crowe JH, Crowe L. M. Preservation of membranes in anhydrobiotic organisms: the role of trehalose. Science 1984 223:701-703.
-
Crowe JH, Clegg JS, Crowe L. M.. Anhydrobiosis: the water replacement hypothesis. In The Roles of Water in Foods. D. S. Reid, editor. Chapman & Hall , New York , 1998 440-455.
-
Donnamaria MC, Howard EI, Grigera JR. Interaction of water with a, a -trehalose in solution: molecular dynamics simulation approach. J. Chem. Soc. Faraday Tra ns. 1994 90: 2731-2735.
-
Cordone L, Cottone G, Giuffrida S, Palazzo G, Venturosi G, Viappiani C. Internal Dynamics and Protein-Matrix Coupling in Trehalose Coated Proteins. Biochim. Biophys. Acta 2005 1749:252-281.
-
Cottone G, Cicciotti G, Cordone L. Protein-trehalose-water structures in trehalose coated carboxy-myoglobin. J. Chem. Phys. 2002 117:9862-9866.
-
Magazù S, Migliardo F, Ramirez-Cuesta AJ. Inelastic Neutron Scattering Study on Bioprotectant Systems. Journal of the Royal Society Interface 2005 2:527-531.
-
Magazù S, Migliardo F, Ramirez-Cuesta AJ. Concentration Dependence of Vibrational Properties of Bioprotectant/Water Mixtures by Inelastic Neutron Scattering. Journal of Royal Society Interface 2007 4:167-172.
-
Colognesi D, Parker SF. The TOSCA Manual, Rutherford Appleton Laboratory UK, 1999.
-
Kolesnikov AI, Li JC, Ahmad NC, Loong C-K, Nipko M, Yocum L, Parker SF. Neutron spectroscopy of high-density amorphous ice. Physica B 1999 263 :650-652.
-
Kolesnikov AI, Li JC, Parker SF, Eccleston RS, Loong C-K. Vibrational dynamics of amorphous ice. Phys. Rev. B 1999 59 :3569-3578.
-
Li JC. Inelastic neutron scattering studies of hydrogen bonding in ices . J. Chem. Phys. 1996 105 :6733-6755.
- Ballone P, Marchi N, Branca C, Magazù S. Structural and Vibrational Properties of Trehalose: A Density Functional Study . J. Phys. Chem. 2000 104 :6313-6317.
AUTHOR:
Dott.ssa Federica Migliardo Biophysics and Soft Condensed Matter Research Group Department of Physics, University of Messina , C.da Papardo Sperone 31 – 98166 Messina ( Italy ) Laboratoire de Dynamique et Structure des Matériaux Moléculaires, U.M.R. n° 8024, Universitè de Lille 1, U.F.R. de Physique – Bât. P5, 59655 Villeneuve D'Ascq Cedex (France)
Keep reading
Biology →
Scoperta causa genetica della sclerosi multipla?
2 June 2016 · 5 min
Scoperta la “via” che permette al calcio di entrare rapidamente negli “orologi della vita” cellulare
20 June 2011 · 3 min