Splicing RNA: "molla molecolare" che risolve paradosso della biologia cellulare

Simulazioni atomistiche del Cnr-Iom di Trieste svelano come la proteina TAT-SF1 tenga in tensione una struttura dell'RNA pronta a scattare per riconoscere le sequenze genetiche corrette.
Introduzione
Ogni volta che una cellula produce una proteina, deve prima "montare" il messaggio genetico di partenza, tagliando le parti inutili e unendo solo quelle che servono. Questo processo si chiama splicing, ed è così preciso che un singolo errore può innescare patologie gravi: alcuni tumori, malattie neurodegenerative, disturbi dello sviluppo.
Il progesso che guida il riconoscimento tra le molecole coinvolte è estremamente complesso, e nessuno capiva come, sebbene la complementarietà chimica tra le sequenze in gioco è bassa, il riconoscimento avvenisse sempre nel posto giusto, in modo rapidissimo. Era un paradosso.
Un nuovo studio pubblicato su Nucleic Acids Research, firmato da ricercatori del Cnr-Iom di Trieste in collaborazione con l'Institute of Cancer Research di Londra, risolve questo enigma. La risposta, osservata per la prima volta a livello atomistico, è elegante: una molecola di RNA viene tenuta in tensione come una molla caricata, pronta a scattare nel momento giusto verso il bersaglio corretto.
Lo splicing: tagliare e ricombinare il messaggio genetico
Il DNA è il manuale di istruzioni della cellula, ma le sue istruzioni non vengono usate direttamente. Prima vengono copiate in una molecola intermedia chiamata RNA messaggero (mRNA, dall'inglese messenger RNA), che poi viene tradotta in proteine. Il problema è che il testo iniziale del messaggio è pieno di sequenze non codificanti chiamate introni, che devono pertanto essere rimosse prima della sintesi delle proteina.
Questo lavoro di "taglia e cuci" è chiamato “splicing”, ed è eseguito da una macchina molecolare di straordinaria complessità chiamata spliceosoma. Al suo interno lavorano centinaia di proteine e cinque molecole di RNA altamente specializzate, le snRNA (dall'inglese small nuclear RNA, piccoli RNA nucleari - sono brevi molecole di RNA non codificante della lunghezza di circa 100-200 nucleotidi).
Ognuna di queste molecole ha un ruolo preciso. Una di esse, chiamata U2 snRNP (dove snRNP sta per small nuclear ribonucleoprotein, piccole ribonucleoproteine nucleari ossia un complesso molecolare formato da RNA non codificante, snRNA e proteine), ha il compito di trovare nell'RNA messaggero una sequenza specifica denominata branch site, che in italiano si può tradurre come "sito di ramificazione".
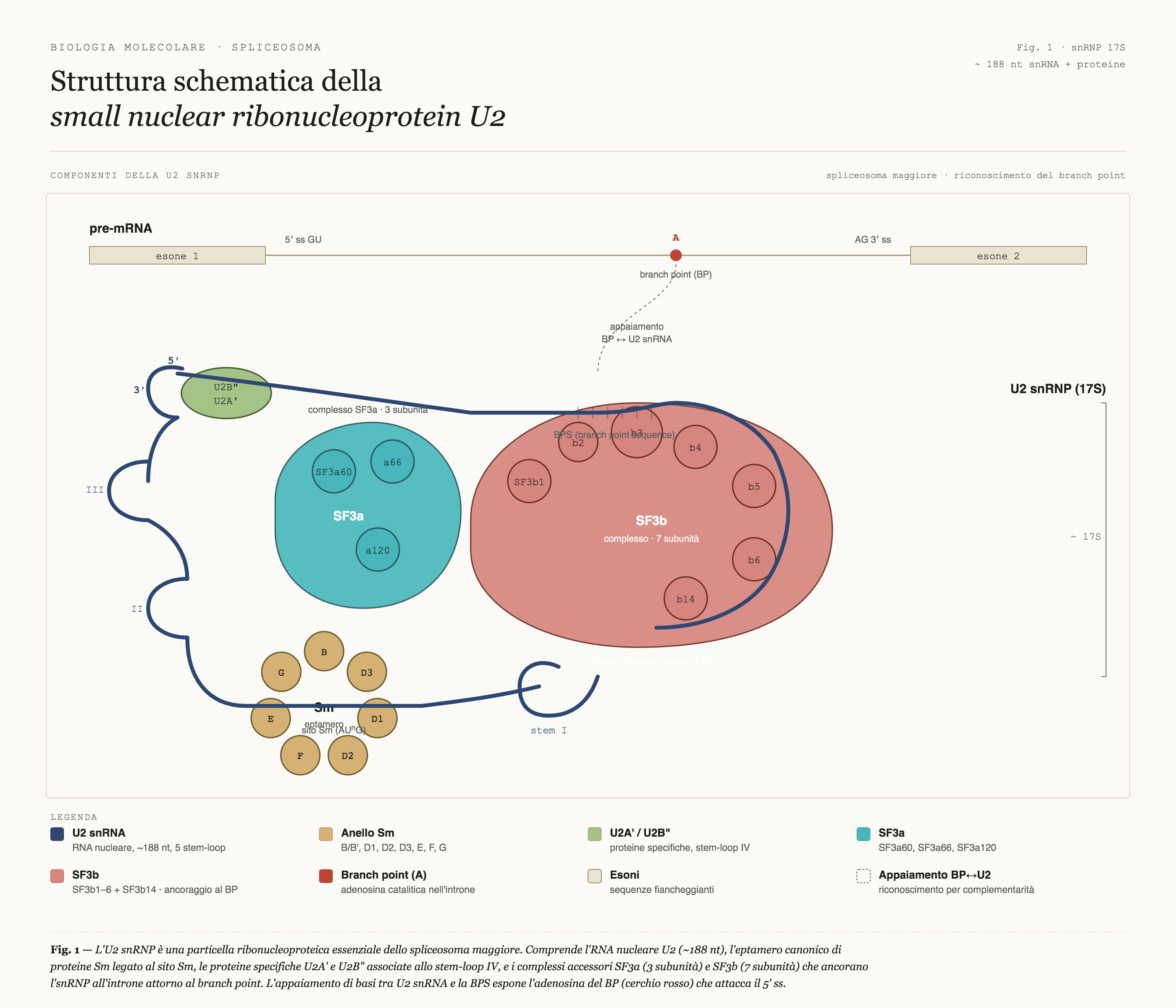
Fig. 1 — L'U2 snRNP è una particella ribonucleoproteica essenziale dello spliceosoma maggiore. Comprende l'RNA nucleare U2 (~188 nt), l'eptamero canonico di proteine Sm legato al sito Sm, le proteine specifiche U2A′ e U2B″ associate allo stem-loop IV, e i complessi accessori SF3a (3 subunità) e SF3b (7 subunità) che ancorano l'snRNP all'introne attorno al branch point. L'appaiamento di basi tra il segmento 5′ di U2 snRNA e la branch point sequence espone l'adenosina del BP (▲) che attacca nucleofilicamente il 5′ ss durante il primo passaggio di splicing.

Fig. 2 il gruppo di ricerca. Pavlína Pokorná (la seconda da sinistra), Alessandra Magistrato (la terza da sinistra in prima fila).
Questo sito funziona come punto di ancoraggio per avviare il taglio dell'introne. Trovarlo è indispensabile. Per anni la domanda rimasta senza risposta è stata: come fa U2 snRNP a trovarlo con tanta precisione, nonostante la scarsa complementarietà tra le sequenze coinvolte? È come avere una chiave che non combacia perfettamente con la serratura: in condizioni normali la porta rimarrebbe chiusa. Eppure la porta si apre sempre. Come è possibile?
La molla caricata: un meccanismo mai osservato prima
Le ricercatrici Alessandra Magistrato e Pavlína Pokorná del Cnr-Iom di Trieste, in collaborazione con Vladimir Pena dell'Institute of Cancer Research di Londra, ha affrontato il problema utilizzando un approccio computazionale. I ricercatori invece di fotografare le molecole, le hanno simulate atomo per atomo.
Le simulazioni hanno rivelato uno stato intermedio che nessuna tecnica sperimentale aveva mai potuto osservare direttamente. Al centro c'è una struttura specifica dell'RNA di U2, chiamata BSL (dall'inglese branch-stem loop): una piccola porzione della molecola ripiegata su se stessa a formare una struttura ad anello nota anche come forcina o ansa forcina.
In condizioni normali, questa struttura è tenuta in una conformazione superavvolta (o superavvolgimento del DNA, il processo tramite cui la molecola a doppia elica si attorciglia ulteriormente su se stessa) e ad alta energia da una proteina chiamata TAT-SF1, che agisce come un chiavistello molecolare. Immaginate un elastico avvolto più volte su se stesso, tenuto fermo da una molletta: non può rilasciarsi, ma ha energia pronta a essere liberata.
Quando avviene il distacco di TAT-SF1 (il distacco è regolato da un'altra proteina, chiamata PRP5) l'elastico si libera. La struttura BSL si distende e, grazie all'energia conformazionale accumulata, invade il filamento dell'RNA messaggero e forma con esso i legami chimici (le cosiddette coppie di basi) che costituiscono il riconoscimento del sito di ramificazione (branch site). Questo processo è definito dai ricercatori "strand invasion", cioè "invasione del filamento".
Il meccanismo è stato chiamato "loaded-spring", molla caricata, proprio perché l'energia non viene generata al momento dell'azione: viene accumulata in anticipo e poi rilasciata nel momento opportuno.
Il paradosso risolto, come funziona il meccanismo nonostante la bassa complementarietà
Qui sta il cuore della scoperta. Normalmente, affinché due molecole di RNA si riconoscano e si leghino, le loro sequenze devono essere chimicamente complementari, come i denti di due ingranaggi che si incastrano. Più sono complementari, più il legame che va a formarsi è spontaneo e stabile.
Nel caso di U2 snRNP e del branch site, questa complementarietà è bassa e variabile da gene a gene. In condizioni ordinarie, l'invasione del filamento non avverrebbe spontaneamente: non ci sarebbe abbastanza "forza" chimica a guidarla.
La molla caricata risolve questo problema in modo elegante: fornisce l'energia che la complementarietà da sola non garantisce, innescando il processo di riconoscimento. Non è la chiave che combacia con la serratura, ma la mano che la spinge la chiave con la forza giusta.
Le simulazioni hanno anche rivelato un dettaglio meccanicistico inatteso: prima che l'invasione del filamento avvenga, il BSL attraversa un ulteriore stato intermedio chiamato "slip-stranded" (filamento scivolato). In questo stato, le coppie di basi interne alla struttura BSL si riorganizzano con uno scorrimento di una posizione, come tessere di un mosaico che si spostano di uno spazio. Questa riorganizzazione libera alcune basi che diventano disponibili per legarsi all'RNA messaggero, avviando l'invasione del filamento. È un passaggio rapidissimo, durato a volte solo pochi nanosecondi nelle simulazioni, ma sufficiente a innescare l'intero processo.
Le simulazioni hanno inoltre dimostrato che l'invasione può procedere in entrambe le direzioni lungo il filamento, raffinando i modelli proposti in precedenza dalla biologia strutturale.
Come si osserva ciò che i microscopi non vedono
Le tecniche classiche di biologia strutturale, come la crioelettromicroscopia (che congela le molecole a temperature criogeniche di circa -196 °C per fotografarle) e la cristallografia a raggi X (che ne determina la struttura attraverso la diffrazione della luce), producono immagini di straordinaria risoluzione. Ma catturano istantanee: la molecola prima del processo e la molecola dopo. Tutto quello che succede nel mezzo rimane, di fatto, invisibile.
Pensate a un acrobata fotografato in piedi al suolo e poi in volo: le due foto esistono, ma il salto, la spinta, la traiettoria (ossia i passaggi intermedi) non compaiono in nessuna delle due immagini.
Le simulazioni della dinamica molecolare fanno esattamente questo: ricostruiscono il salto nel corso del tempo. Calcolano, passo dopo passo, come ogni atomo si muove nel tempo in risposta alle forze degli atomi vicini. Il prezzo da pagare è la potenza di calcolo richiesta, che in questo studio è stata fornita dal supercomputer del CINECA, il principale centro di calcolo italiano, tramite l'iniziativa ISCRA (Italian SuperComputing Resource Allocation).
In tutto questo il vantaggio reale emerge però dall'integrazione dei due approcci: i dati sperimentali della crioelettromicroscopia forniscono i punti di partenza e di arrivo, le simulazioni ricostruiscono i passaggi intermedi. I due metodi non si sostituiscono, si completano a vicenda.
«L'integrazione tra biologia strutturale e simulazioni computazionali avanzate», ha spiegato Magistrato, dirigente di ricerca Cnr-Iom presso la SISSA (Scuola Internazionale Superiore di Studi Avanzati) di Trieste, «ci consente di capire come funzionano sistemi biologici estremamente complessi e dinamici con un livello di accuratezza che nessuno dei due approcci da solo potrebbe raggiungere.»
Implicazioni cliniche: dallo splicing ai tumori
Lo splicing non è biologia astratta. Errori in tale meccanismo sono collegati all’insorgenza di numerose malattie. Alcune forme di cancro sfruttano difetti nello splicing per produrre versioni alterate di proteine che normalmente controllano la crescita cellulare. Malattie come la SMA (atrofia muscolare spinale, una malattia neuromuscolare genetica), in alcune sue forme, derivano direttamente da mutazioni che compromettono il riconoscimento del branch site, esattamente il passaggio che questo studio ha chiarito per la prima volta con un dettaglio di livello atomico.
Conoscere come funziona la molla caricata significa avere una mappa più accurata di un processo che, quando si rompe, può innescare la malattia. Non si tratta ancora di una terapia, ma di una maggiore comprensione del meccanismo che potrebbe rendere possibile, in prospettiva, la progettazione di molecole capaci di intervenire in modo mirato. L’intervento, su cui il gruppo della dott.ssa Magistrato lavora da anni consiste nel bloccare il processo di riconoscimento in modo selettivo in cellule tumorali che dipendono da uno splicing alterato.
Il lavoro è stato finanziato dall'AIRC (Associazione Italiana per la Ricerca sul Cancro) con un progetto Investigator Grant (si tratta di un finanziamento di ricerca assegnato a scienziati esperti e indipendenti (detti Principal Investigator) per condurre un progetto scientifico specifico, a conferma dell'interesse clinico della ricerca.
Un paradigma che potrebbe valere oltre lo splicing
In tutto il ragionamento fatto finora c'è un aspetto della scoperta che i ricercatori che hanno condotto lo studio sottolineano come potenzialmente più ampio, il meccanismo della molla caricata potrebbe non essere caratteristico solo dello splicing.
L’articolo scientifico suggerisce esplicitamente che questo principio, una struttura di RNA mantenuta in tensione da una proteina e poi rilasciata per guidare un riconoscimento molecolare, potrebbe rappresentare un paradigma generale per il riconoscimento di acidi nucleici in altri contesti biologici. In altre parole: ovunque nella cellula ci sia bisogno di riconoscere una sequenza con precisione nonostante una bassa complementarietà, potrebbero esistere meccanismi analoghi ancora non scoperti.
«Il nostro approccio modellistico», ha spiegato Pokorná, prima autrice dello studio, «può fungere da guida per studi analoghi su altri processi cellulari, aprendo nuove finestre su come funziona l'espressione genica.»
Conclusioni
Lo splicing dell'RNA è uno dei processi cellulari più importanti e meno visibili. Questo studio dimostra che le molecole protagoniste non si limitano ad aspettare che le condizioni siano favorevoli: vengono preparate, indirizzate verso il bersaglio con una logica fisica precisa.
Il paradosso è risolto: la bassa complementarietà non è un ostacolo, perché la cellula lo aggira con un meccanismo attivo. E ora che il meccanismo è stato compreso è lecito porsi altre domande. Quante altre "molle caricate" stanno ancora aspettando di essere scoperte nel paesaggio molecolare della cellula? E scoprirle che cosa significherebbe per quanto riguarda la comprensione e il trattamento delle malattie che nascono quando questo paesaggio si guasta?
Fonti
Pavlína Pokorná, Vladimir Pena, Alessandra Magistrato, U2 snRNP recognizes the branch site through a loaded-spring strand-invasion mechanism, Nucleic Acids Research, Volume 54, Issue 9, 22 maggio 2026, gkag429.